Внешний вид эктотрофной микоризы дуба
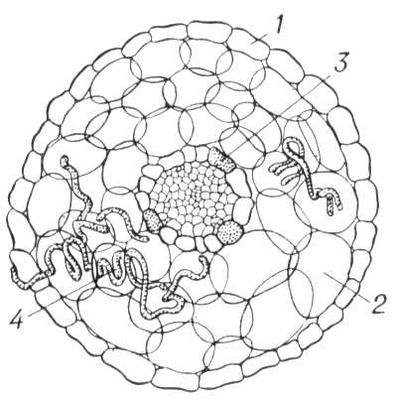
Поперечный разрез эндотрофной микоризы клёна:
1 — эпидермис;
2 — кора;
3 — эндодерма;
4 — клубки гиф гриба.
Симбиозом (от греческого "symbiosis" - "совместная жизнь") в биологии принято называть такое взаимодействие двух и более разных организмов, от которого все партнеры получают пользу, все что-то выигрывают. В широком смысле симбиоз охватывает все формы тесного сожительства организмов разных видов, включая и паразитизм, который в этом случае называется антагонистическим симбиозом.
Обычно симбиоз бывает мутуалистическим, т. е. сожительство обоих организмов (симбионтов) взаимовыгодно и возникает в процессе эволюции как одна из форм приспособления к условиям существования. Симбиоз может осуществляться как на уровне многоклеточных организмов, так и на уровне отдельных клеток (внутриклеточный симбиоз). В симбиотические отношения могут вступать растения с растениями, растения с животными, животные с животными, растения и животные с микроорганизмами, микроорганизмы с микроорганизмами. Термин «симбиоз» впервые введён немецким ботаником А. де Бари (1879) в применении к лишайникам. Яркий пример симбиоза среди растений представляет микориза — сожительство мицелия гриба с корнями высшего растения (гифы оплетают корни и способствуют поступлению в них воды и минеральных веществ из почвы); некоторые орхидеи не могут расти без микоризы.
|
Внешний вид эктотрофной микоризы дуба |
|
Поперечный разрез эндотрофной микоризы клёна: 1 — эпидермис; 2 — кора; 3 — эндодерма; 4 — клубки гиф гриба.
|
|---|
Примеры симбиоза животных и растений — сожительство одноклеточных водорослей с разными животными — простейшими, кишечно-полостными (гидры, коралловые полипы), ресничными червями и др. Показано, что одноклеточные водоросли, поселяющиеся в клетках коралловых полипов, играют важную роль в нормальном росте и развитии последних.
 |
|
|---|
Широко известен пример симбиоза между раками-отшельниками и актиниями. Последние поселяются на раковине, в которую прячет своё брюшко рак-отшельник. Стрекательные клетки щупалец актиний — надёжная защита обоих симбионтов. Питается актиния за счёт остатков пищи, активно добываемой раком.
В самом простом варианте рак Eupagurus excavatus ищет пустую раковину с уже прикрепившейся к ней актинией. В случае удачи он переползает из своей раковины в найденную.
При более развитых симбиотических отношениях другого рака-отшельника Pagurus arrosor с актинией рак не ищет пустую раковину с анемоной. Он может снять ее с любого субстрата и пересадить на свой домик. Когда рак-отшельник вырастает и переходит в другую, большую, раковину, он пересаживает на нее и свою актинию. При прикосновении клешни рака к актинии она вначале начинает сжиматься. Но, удивительно, на дальнейшее похлопывание клешней актиния не отвечает ни выбрасыванием стрекательных нитей, ни продолжением сокращения тела. Напротив, она распускается. Даже полностью сократившаяся актиния вновь распускается при поглаживании раком-отшельником. Затем рак начинает поглаживать подошву «цветка», после чего она сокращается и отделяется от субстрата. Раку остается лишь пересадить актинию на свой новый домик. Иногда, правда, морская красавица и сама переходит на новое место жительства, вначале наклоняясь и охватывая раковину щупальцами, а затем переворачиваясь и прикрепляясь к ней подошвой. Если на раковине остается достаточно места, предприимчивый рак может обзавестись еще одной актинией. Описаны случаи, когда Parrosor носил на своей раковине до 8 актиний.
Однако наиболее совершенный симбиоз наблюдается у рака-отшельника Eupagurus pricle-axi и актинии Aclainsia palliata. В этом случае рак захватывает еще молодую актинию и больше не расстается с ней. Правда, иногда рак-отшельник нарушает свою верность, уступив в сражении более сильному сородичу и раковину, и актинию. Смена домика не обязательна для этого рака в отличие от других раков благодаря актинии. Ее подошва обрастает всю раковину, окольцовывая устье. Если на раковине находятся две актинии, то их подошвы смыкаются, также охватывая всю раковину. Подошва Адамсии выделяет плотную слизистую пластину, которая быстро твердеет. По мере роста актинии эта пластина перерастает край устья раковины и нависает над телом рака. В результате домик постоянно увеличивается и раку не приходится искать более просторное жилище. Этот домик намного легче толстостенной раковины, эластичен и не затрудняет движений рака.
Актинии, однако, не всегда являются пассивной стороной при создании симбиотических пар. Например, Autholoba retic-ulata сама ищет партнера. Поселяясь на камнях, она держится за субстрат щупальцами, а не подошвой. В таком положении актиния ожидает рака, при появлении которого ее подошва схватывает его лапку. После этого вся анемона переползает на панцирь рака.
Все эти типы отношений раков-отшельников с актиниями являются истинным симбиозом, потому что выгодны обоим партнерам: рак защищен от врагов стрекательными нитями актиний, у актинии же улучшаются условия питания как за счет расширения зоны охоты, так и за счет «стола» рака-отшельника.

Широко распространён симбиоз животных (и человека) с микроорганизмами, например образующими нормальную кишечную флору . У некоторых насекомых переваривание клетчатки осуществляется ферментами, выделяемыми дрожжевыми клетками, живущими в их пищеварительном тракте, в особых углублениях кишечника. Микробы заселяют кишечник хозяина с первых часов жизни, попадая туда главным образом с пищей. В период младенчества люди обладают высоким содержанием бифидобактерий. У здорового ребенка первого года жизни 90-98% всего микробиоциноза толстого кишечника составляет бифидофлора.
Бифидобактерии - микроорганизмы, которые заселяют нижние отделы кишечника и выполняют следующие функции:
С развитием организма человека (от новорождённого до взрослого состояния) кишечная флора меняется, пока не приобретёт определённую стабильность — так называемая нормальная кишечная флора, состоящая из 2 групп микроорганизмов: 1) комменсальные формы ( Комменсал и зм, сотрапезничество, нахлебничество, сожительство животных разных видов, характеризующееся тем, что один из них (комменсал) постоянно или временно живёт за счёт другого, не причиняя ему вреда) и сапрофиты ( Сапрофиты (от греч. Sapros — гнилой и phyton — растение, растения, питающиеся органическими веществами отмерших организмов или выделениями живых) , 2) условно, или потенциально, болезнетворные.
В верхнем отделе тонких кишок микробов сравнительно мало, т. к. большинство их погибает в желудке под действием соляной кислоты желудочного сока. Здесь преобладают аэробные стрептококки (так называемые энтерококки), лактобациллы и дрожжи. В нижних отделах кишечника больше грамотрицательных бактерий, главным образом из группы кишечной палочки, и спороносных бацилл. В 1 г кала содержится 3—5·1011 микробов, а в содержимом всего кишечника — около 1015 микроорганизмов. Нарушение видового состава и количественных соотношений разных микробов в кишечной флоре, вызываемое, например, антибиотиками, называется дисбактериозом (или дисбиозом). От кишечной флоры, её состава и состояния зависят течение процессов пищеварения, образование ряда ферментов (например, целлюлазы, разлагающей клетчатку) и других физиологически активных веществ (аминокислот, нуклеотидов, витаминов), не синтезируемых организмом хозяина.
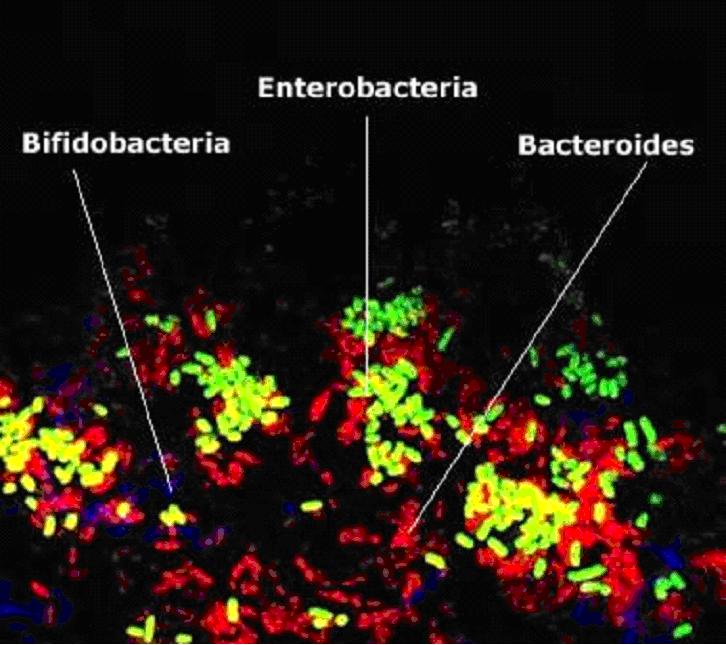
На рубеже ХХI века сформировалось представление о микрофлоре организма человека как о еще одном органе, покрывающем в виде чулка кишечную стенку, другие слизистые оболочки и кожу человека. Оставаясь невидимым, этот «орган» весит около двух килограммов и насчитывает порядка 1014 сто биллионов) клеток микроорганизмов. Это число в десять раз превышает число собственных клеток организма-хозяина, то есть - человеческих.
 |
Для бобовых растений важное значение имеет симбиоз с клубеньковыми бактериями. Есть почвенные бактерии, которые способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Это азотофиксирующие бактерии. Они живут самостоятельно или поселяются в корнях растений. После проникновения в корни, эти бактерии стимулируют деление клеток, ткани корней разрастаются и на них образуются клубеньки. Такие бактерии называют клубеньковыми. Растения используют азотные соединения, которые выделяют эти бактерии. В свою очередь от растений бактерии получают углеводы и минеральные соли. Таким образом тесная связь, возникающая между зелеными растениями и клубеньковыми бактериями, оказывается взаимно полезной. |
|---|
Многочисленны и разнообразны формы симбиоза простейших с различными прокариотными организмами (бактериями, риккетсиями и др.), локализующимися обычно в цитоплазме. Известны случаи симбиоза инфузорий с бактериями, поселяющимися в ядре простейшего.
В 1914 г. датское исследовательское судно "Сибога" во время глубоководных тралений вблизи Зондского архипелага добыло несколько экземпляров невзрачных морских червеобразных организмов, которые обитали в длинных тонких трубочках. В честь экспедиционного судна им было дано название Siboglinum. Эти организмы сначала не вызвали большого интереса среди биологов, поскольку были отнесены к обычным в морских сообществах многощетинковым червям-полихетам. Последующие морские экспедиции приносили новые находки подобных животных, и постепенно, зоологи стали осознавать, что эти организмы сильно отличаются от многощетинковых червей. Их таксономических ранг стал постепенно повышаться, сначала до отдельного отряда, потом до класса, и, наконец, в 1944 г. профессор Московского университета В.Н.Беклемишев выделил их в отдельный тип - самую крупную категорию в систематике животного царства. Название этого типа Pogonophora происходит от греческих слов pogon - борода и phorоs - носитель (на переднем конце у погонофор имеется венчик длинных извитых щупалец, напоминающих бороду). Глубоководные траления, проведенные во время экспедиций советского исследовательского судна "Витязь", принесли множество видов погонофор из всех районов мирового океана. Обработкой этих богатейших коллекций занимался профессор Ленинградского университета, впоследствии академик А.В.Иванов, который внес решающий вклад в изучение систематики, строения и развития погонофор.
Самая загадочная черта организации погонофор состояла в том что, эти организмы не имели ни рта, ни кишечника, и, следовательно, способ питания погонофор оставался загадкой. Для объяснения этого факта предлагались самые невероятные объяснения. Предполагалось, например, что погонофоры удовлетворяют все потребности питания за счет всасывания растворенных органических веществ в следовых количествах имеющихся в морской воде. В связи с таким необычным способом питания предполагалось, что погонофоры обладают крайне низким уровнем обмена и, стало быть, крайне медленным ростом и большой продолжительностью жизни. Как это ни покажется странным сейчас, в 50-60-ые годы всерьез обсуждался вопрос о продолжительности жизни погонофор в 40-50 тыс. лет. Дело в том, что трубочки погонофор воткнуты в грунт почти вертикально и пронизывают слои осадка, образовавшиеся десятки тысяч лет назад. До начала 60-ых годов двадцатого века никаких органов, позволяющих погонофорам закапываться в грунт, найдено не было. Предполагалось, что личинки погонофор оседали на поверхность осадка, а затем медленно, в течение нескольких десятков лет росли вместе с накоплением слоев осадка. Это фантастическое предположение было отвергнуто после того, как на заднем конце тела погонофор был найден особый сегментированный хвостик, действующий как маленькая лопата и закапывающий погонофору в грунт по мере роста.
Другое предположение, выдвинутое А.В.Ивановым, состояло в том, что погонофоры обладают наружным пищеварением в бокале щупалец. Наличие на щупальцах тончайших одноклеточных выростов - пиннул, снабженных кровеносными капиллярами, рассматривалось как косвенное подтверждение возможности всасывания пищи через покровы. Делались безуспешные попытки найти в щупальцах пищеварительные энзимы. Оставалось непонятным и то, а почему, собственно, погонофоры прибегли к такому странному способу питания, редуцировав рот и кишечник, которыми пользуются все остальные животные. Тем не менее, эта гипотеза была принята многими авторами и до начала 80-ых годов двадцатого века излагалась в учебниках по зоологии.
Новый этап в изучении погонофор начался со второй половины 70-ых годов, когда в так называемых рифтовых зонах океана были найдены представители новой группы погонофор - вестиментиферы. Рифтовые зоны - это система трещин, возникших в участках стыка литосферных плит, из которых состоит верхняя мантия Земли. В таких местах сквозь толщу океанической коры просачиваются горячие газы, нагревающие воду до температуры 300-400 градусов (при высоком давлении на больших глубинах вода не кипит даже при температуре в несколько сот градусов). В этой воде растворено много сероводорода и сульфидов металлов (железа, цинка, никеля, меди), которые окрашивают ее в черный цвет. Концентрации металлов в горячей воде в этих местах превышают таковые в обычной морской воде в 100 млн. раз. Струи этого горячего раствора смешиваются с холодной водой придонных слоев океана, охлаждаются, сульфиды выпадают в осадок и формируют особые конические постройки высотой несколько десятков метров - черные курильщики. Так их называют из-за мощных потоков горячего сульфидного раствора, напоминающих клубы черного дыма.
Само собой разумеется, что подобные районы всегда были интересны для геологов, так как позволяли воочию наблюдать, как формировались ценнейшие сульфидные руды. Правда, целенаправленные исследования таких гидротермальных оазисов стали проводить только после создания специальных глубоководных управляемых аппаратов - миниатюрных подводных лодок с небольшим экипажем исследователей, способных погружаться на глубины в несколько тысяч метров и собирать образцы грунта с помощью механических манипуляторов.
Чего не ожидали исследователи, так это - наличия в рифтовых зонах богатой фауны. На больших глубинах океана, куда никогда не проникает солнечный свет и где вся фауна питается остатками отмерших организмов, падающих из богатых жизнью верхних слоев воды, численность и биомасса животных очень малы. Рифтовые зоны с их горячими вулканическими газами, содержащими большие концентрации сероводорода, тяжелых металлов и других ядовитых для большинства организмов соединений, казалось, должны быть долинами смерти среди и так не слишком богатых жизнью морских глубин. Однако, первые же фотографии, сделанные исследователями через иллюминаторы подводных аппаратов, показали колоссальное обилие живых существ в составе этих гидротермальных сообществ.
 |
 |
|---|
На снимках было видно, что склоны черных курильщиков почти до самых вершин покрыты толстым слоем бактерий (сплетения миллиардов бактериальных клеток образуют так называемые маты), способных выживать при температуре до 120 градусов. В отдалении от устья курильщиков, там где температура опускается ниже 40 градусов, на уступах курильщиков были видны сплетения белых трубок гигантских (до 2, 5 м) червей с ярко алыми щупальцами. В зарослях трубок ползали крабы, рядом плавали рыбы, в расселинах сидели крупные (20-30 см) двустворчатые моллюски, попадались осьминоги, словом, жизнь кипела. Красота и богатство сообществ черных курильщиков, резко контрастирующие с бедным и однообразным населением ложа океана, так поразили исследователей, что некоторые из гидротермальных оазисов называются в научной литературе весьма поэтично: "Райский сад", "Розовый сад" и т.п.
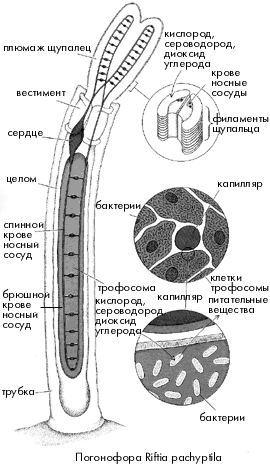
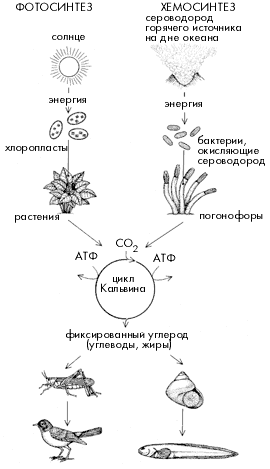
У вестиментифер (так же как у погонофор) во взрослом состоянии нет рта и кишечника. По оси туловищного отдела вестиментифер проходит массивный клеточный тяж, который сначала считался запасающим органом и был назван "трофосома". Электронно-микроскопические исследования показали, что крупные клетки трофосомы содержат множество вакуолей с бактериями. Бактерии вестиментифер принадлежат к группе сероводородокисляющих бактерий. Они окисляют сероводород до серы (а потом до серной кислоты, нейтрализуемой карбонатами), и полученную при этом энергию используют для синтеза органических веществ из углекислого газа и воды. Этот процесс носит название хемосинтеза и характерен для многих видов свободноживущих бактерий, обитающих там, где в окружающей среде много сероводорода и есть кислород.
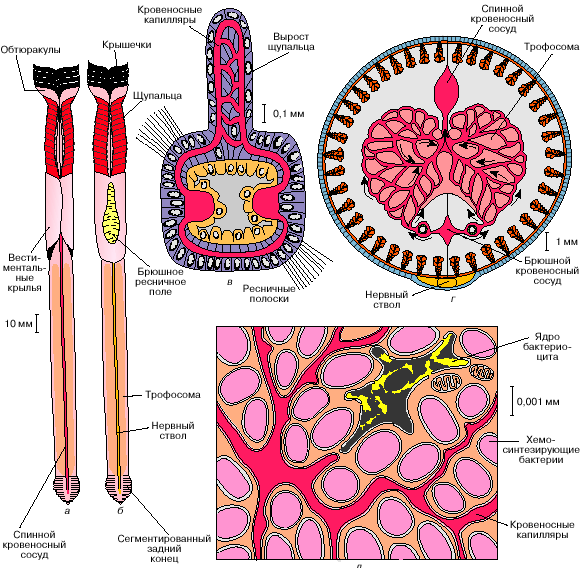
В гидротермальных оазисах сероводород поступает из черных курильщиков, а кислород - за счет подсоса холодной и богатой кислородом глубинной воды, окружающей зоны гидротермальных источников. Сероводород – очень ядовитое вещество, практически у всех животных он блокирует дыхание, занимая места связывания кислорода на молекулах гемоглобина и инактивируя важный дыхательный фермент – цитохром-с-оксидазу.
Однако вестиментиферы отлично живут при таких концентрациях сульфида в окружающей среде, которые для большинства живых организмов смертельны. Мало того, их ярко-красные щупальца-жабры улавливают в воде и передают в кровь (на долю крови у этих животных приходится более 30% общего объема тела) одновременно и кислород, нужный для дыхания самих погонофор, и сероводород, необходимый питающим их бактериям. Исследования показали, что гемоглобин вестиментифер совершенно не похож на гемоглобин большинства других живых существ. Его молекула очень крупная (молекулярная масса составляет 2 млн дальтон (Да), в то время как молекулярная масса гемоглобина человека – 64000 Да). При этом гемоглобин вестиментифер содержится в плазме крови в свободном виде, а не заключен внутри эритроцитов, как у позвоночных животных. Гемоглобин вестиментифер способен одновременно связывать и кислород, и сероводород – молекулы этих веществ присоединяются к большой молекуле гемоглобина в разных участках. Таким образом решаются сразу две задачи – сульфид не нарушает дыхание животного и при этом не окисляется кислородом до попадания в бактериальную клетку. Сложная кровеносная система вестиментифер содержит две системы капилляров: одну в щупальцах, а другую в трофосоме. Сеть капилляров кровеносной системы проникает непосредственно внутрь клеток трофосомы и при этом так густа, что любую бактерию от ближайшего капилляра отделяет не более двух других бактерий. Гемоглобин вестиментифер соединяется и с кислородом и с сероводородом, при этом сероводород обратимо связывается с белковой частью молекулы, а кислород - с гемом. Впрочем, при недостатке кислорода бактерии способны получать его, переводя нитраты, которыми богаты глубинные воды океана, в нитриты. Бактерии, защищенные внутри организма хозяина от неблагоприятных воздействий, получают от него сероводород и кислород. За счет самопереваривания части клеток трофосомы вместе с бактериями хозяин получает органические вещества, которые служат единственным источником питания вестиментифер. Таким образом, сожительство хемосинтезирующих бактерий и вестиментифер является взаимовыгодным симбиозом.
Хотя вестиментиферы являются преобладающими по численности многоклеточными, живущими здесь, симбиоз с хемотрофными бактериями свойствен не только им. Исследования показали, что подобным же образом получают пищу и двустворчатые моллюски двух обитающих около гидротермальных выходов видов – Calyptogena magnifica и Bathymodiolus termophilus. У Calyptogena серобактерии поселяются на жабрах, где они могут легко получать необходимые им кислород и углекислый газ. Однако сероводорода в окружающей жабры моллюска воде относительно немного – в отличие от рифтий, гемоглобин Calyptogena необратимо инактивируется сульфидом. Так, чтобы иметь возможность и есть, и дышать, моллюскам приходится располагаться на границе чистой воды и исходящих из источника струй, насыщенных химическими соединениями. В такую струю Calyptogena опускает свою далеко вытягивающуюся ногу. В крови моллюска присутствует особый транспортный белок, способный лучше гемоглобина и цитохром-с-оксидазы связывать сероводород и тем самым предотвращающий блокирование дыхания. С помощью этого белка сульфид и переносится с током крови от места поглощения к бактериальным клеткам на жабрах, не окисляясь и не отравляя по дороге самого моллюска.
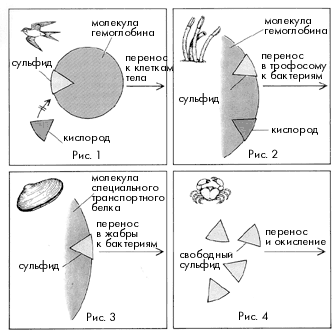
У представителей второго вида двустворчатых, Bathymodiolus thermophilus, особенности транспорта H 2 S не изучены, известно только, что и у них серобактерии поселяются на жабрах. Оба вида моллюсков, по-видимому, уже не способны питаться самостоятельно, без помощи симбионтов, и на тех участках дна, где выход содержащих сероводород горячих источников прекратился, эти животные погибают.
Остальные многоклеточные обитатели гидротермалей, во всяком случае, крабы, креветки и рыбы, питаются обычным образом. Они либо отфильтровывают мелкие пищевые частицы (в том числе и бактерий) из воды, либо хищничают, обкусывая, например щупальца вестиментифер. Однако и таким животным приходится вырабатывать определенные приспособления для жизни в столь специфических условиях. Крабы, например, способны нейтрализовать ядовитый сероводород, окисляя его до менее токсичного тиосульфата. Происходит этот процесс в гепатопанкреасе – специальной ткани, по своим функциям сходной с печенью позвоночных животных.
В общем, подводные «райские сады» являются удивительными, совершенно необычными экосистемами, механизмы адаптации членов которых изучены еще далеко не полностью.
Открытие симбиотрофного (обеспечиваемого симбионтами) питания у вестиментифер, натолкнуло исследователей на мысль, что таким же способом могут питаться и типичные погонофоры, известные задолго до открытия гидротермальных оазисов. В их организме имеется загадочный орган - замкнутый с обоих концов срединный канал. В клетках этого органа были найдены бактерии, что позволило считать срединный канал гомологом трофосомы. Правда, бактерии, найденные у погонофор принадлежат к другой группе прокариотных организмов - это метанокисляющие бактерии. Они окисляют метан и за счет полученной энергии синтезируют органическое вещество.
Откуда же берется метан в толще грунта? Оказывается, что в таких высоких концентрациях (около 1 мл на кубический дециметр грунта), при которых бактерии способны не только существовать, но еще и "кормить" хозяина, метан может появиться, прежде всего, в результате просачиваний из подводных месторождений нефти и газа. Поэтому места обитания погонофор перспективны для поисков подводных залежей этих ценнейших ископаемых. Интересно, что те районы, в которых обитают немногие относительно мелководные виды погонофор (Северное море, прибрежные районы вблизи о. Сахалин, Баренцево море) - это как раз районы в которых уже ведется добыча нефти и газа или известны их запасы.
Большинство представителей класса погонофор - обитатели больших глубин Мирового Океана, где пока нефть и газ не добывают и даже не ищут. Современные технологии пока не рассчитаны на добычу полезных ископаемых с больших глубин. Но недалеко то время, когда мелководные месторождения истощатся. Вот тогда погонофоры и укажут нам, где нужно искать нефть и газ на больших глубинах.
Исследования яйцеклеток вестиментифер показало, что бактерий в них нет и, следовательно, бактериальные симбионты от матери к потомству не передаются. Откуда же берутся бактерии, живущие в клетках трофосомы вестиментифер?
Ответ на этот вопрос удалось получить в результате изучения личиночного развития вестиментифер. Оказалось, что личинки вестиментифер имеют нормально развитый рот и кишечник. В течение нескольких суток они плавают в толще воды с помощью венчика ресничек, затем опускаются на субстрат и ползают по поверхности грунта. Они заглатывают хемосинтезирующих бактерий из внешней среды, заражаются ими, после чего рот и анус у молодых вестиментифер редуцируются, а кишечник превращается в орган бактериального питания - трофосому. Недавние исследования показали, что личинки типичных погонофор тоже имеют нормальный рот и кишечник и заражаются симбионтами (метанокисляющими бактериями) из внешней среды. Любопытно, что трофосома погонофор сохраняет просвет (недаром же ее раньше называли срединным каналом) и больше похожа на кишечник, чем сильно видоизмененная трофосома вестиментифер.
Органическое вещество в обычных сообществах происходит за счет процесса фотосинтеза и далее мигрирует по пищевым цепям, пока не подвергнется распаду в организмах животных, грибов и гетеротрофных бактерий. Органическое вещество в большинстве глубоководных сообществ также имеет фотосинтетическое происхождение. Когда-то оно было произведено планктонными водорослями в поверхностном слое воды толщиной 50-100 м. Эти водоросли были съедены планктонными ракообразными, которые стали пищей планктоядным рыбам, а те, в свою очередь, стали жертвами хищных рыб, кальмаров и зубатых китов. Не полностью разложившиеся остатки организмов, погибших в верхних слоях воды, опускаются в глубины океана и становятся пищей для немногочисленного донного населения. Разумеется, до дна доходит лишь ничтожная доля органического вещества, произведенного в верхних слоях воды: ведь пока трупик рачка или мертвое тело крупной акулы пройдет свой "последний" путь длиной от 3 до 6 км по вертикали (а над океанскими впадинами этот путь удлиняется до 11 км), он может быть несколько раз съеден разнообразными организмами, населяющими толщу воды (не говоря уже о бактериях, которые на всем пути продолжают свою разрушительную работу). Вот почему биомасса донных организмов на больших глубинах океана составляет всего 0,1 - 0,2 г на кв. м.
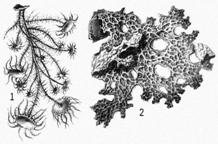
В гидротермальных оазисах были найдены представители некоторых групп животных, считавшихся вымершими более сотни миллионов лет назад. Такие формы обычно называют "живыми ископаемыми". К их числу принадлежат сидячие усоногие ракообразные из рода Неолепас (Neolepas). Усоногие очень обычны на малых глубинах Мирового океана (к ним принадлежат, например, морские желуди и морские уточки, поселяющиеся на камнях, сваях и днищах судов). Они питаются, отфильтровывая из морской воды мелкие пищевые частицы (обычно это клетки планктонных водорослей). Неолепасы питаются точно также: они отфильтровывают из толщи воды сгустки хемосинтезирующих бактерий. Интерес к роду Неолепас связан с тем, что его представители характеризуются особенностями строения, которые не встречаются среди современных усоногих раков. Зато Неолепасы удивительно напоминают ископаемых усоногих, обитавших на мелководьях морей 230-130 млн. лет назад и считавшихся вымершими. Вероятно, гидротермальные оазисы стали убежищами для этих архаичных форм, не сумевших выдержать на мелководьях конкуренцию с более молодыми видами.
Что касается вестиментифер, то после открытия их в гидротермальных оазисах, геологи нашли объяснения находкам окаменевших трубок, которые встречаются в полиметаллических рудах. Трубки вестиментифер известны в залежах цинковых, медных и серебряных руд, образовавшихся в раннем каменноугольном периоде, то есть около 350 млн. лет назад. Таким образом, вестиментиферы - также принадлежат к древнейшим представителям фауны гидротермальных оазисов.
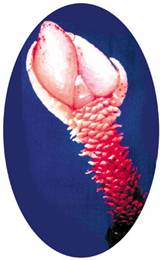 |
Перед нами «живое ископаемое» – такие формы жили больше 130 миллионов лет назад. Этот рачок фильтрует бактерий, как и его родственники – морские желуди. Свое название «неолепас зевинус» он получил в честь известного российского зоолога Г. В. Зевиной |
|---|
Открытие гидротермальных “оазисов”, поддерживаемых энергией хемосинтеза и метанокисления, породило представление об энергетической “независимости” гидротермальных систем от продукции фотосинтеза и, следовательно, от солнечной энергии. Для биосферы Земли такая ситуация была бы уникальной. О полной изоляции, правда, речь не шла, т.к. всем гидротермальным организмам, в том числе, аэробным хемосинтезирующим и метанокисляющим бактериям, требуется кислород, представляющий собой продукт фотосинтеза. Тем не менее, представление об относительной независимости обитателей гидротерм от солнечной энергии сохранилось и по сей день.
Несмотря на ключевую роль хемосинтетической продукции в функционировании большинства глубоководных гидротермальных сообществ, количественная сторона этой “роли” остается не изученной. По сути, лишь вестиментиферы и первиатные погонофоры, во взрослом состоянии полностью лишенные пищеварительного тракта, целиком зависят от симбионтов. Большинство же прочих симбиотрофных животных сочетают симбиотрофию с голозойным питанием (путем сестонофагии или детритофагии). Доля хемосинтетической органики при этом остается неизвестной. Одна из первых попыток оценить этот вклад сделана для креветок Rimicaris exoculata. Так, согласно расчетам авторов, использовавших данные о соотношении стабильных изотопов углерода и метод “баланса масс”, за счет эктосимбионтов взрослые креветки получают более 80% углерода, а молодь около 30%.
Так или иначе, продукция фотосинтетического происхождения играет определенную роль в энергетике гидротермальных экосистем. Эта роль может оказаться существенной, если учитывать, что планктонным личинкам многих гидротермальных животных, в том числе и вестиментифер, приходится питаться продукцией фотосинтеза вне гидротермальных сообществ. Так, недавно выяснилось, что креветки R. exoculata на личиночной (планктонной) стадии жизненного цикла накапливают огромное количество липидов, содержащих полунасыщенные жирные кислоты фотосинтетического происхождения. Липиды служат запасным питательным веществом, обеспечивающим планктонные стадии креветок резервом пищи при поиске гидротермальных биотопов (это в той или иной степени оказалось свойственно всем гидротермальным креветкам. При попадании на активное поле креветки приобретают симбионтов и переходят к симбиотрофному способу питания. Липидный резерв при этом начинает постепенно расходоваться. У взрослых особей липиды обнаруживаются лишь в незначительном количестве. Интересно, что липиды фотосинтетического происхождения были обнаружены также у вестиментиферы Ridgea piscesae. Их происхождение пока не понятно, но возможность накопления этих липидов на личиночной стадии вполне вероятна. Весьма вероятно, что и прочие представители гидротермальной фауны, в том числе, специализированные (особенно их личиночные стадии), в большей или меньшей степени зависят от продукции фотосинтетического происхождения, и это накладывает серьезные ограничения на представления об энергетической независимости гидротермальных экосистем, в том числе и в прошлые геологические эпохи.
Немного по-другому складываются отношения при паразитизме. Паразитизм, форма взаимоотношений между организмами (растениями, животными, микроорганизмами), относящимися к разным видам, из которых один (паразит) использует другого (хозяина) в качестве среды обитания и источника пищи, возлагая при этом (частично или полностью) на хозяина регуляцию своих отношений с внешней средой. Паразитизм — преимущественно экологическое понятие, аналогичное понятиям планктонного, бентосного, почвенного и им подобного образа жизни организмов. Качественная особенность паразитизма, по сравнению с другими формами существования организмов, определяется своеобразием среды обитания, которой для паразита является другой живой организм (хозяин), активно реагирующий на присутствие паразита. Внешняя среда паразита имеет двойственный характер. Различают среду 1-го порядка — окружающие паразита органы и ткани хозяина — и среду 2-го порядка, окружающую хозяина. Взаимоотношения паразита со средою 2-го порядка в основном регулируются через хозяина, хотя имеет место (в различной степени для разных форм паразитов) и прямое воздействие факторов внешней среды (например, температуры) на паразита. Между паразитом и хозяином устанавливаются более или менее глубокие метаболические связи. Многие паразиты являются антигенами , вызывая образование в организме хозяина антител , что, в свою очередь, приводит к реакциям иммунитета . Различают внешний (наружный) паразитизм, или эктопаразитизм (паразит живёт на поверхности тела хозяина), и внутренний паразитизм, или эндопаразитизм (паразит живёт в теле хозяина). В зависимости от продолжительности паразитирования различают временный паразитизм и стационарный паразитизм, включающий периодический и постоянный паразитизм.
Пути возникновения паразитизма в эволюции разнообразны. Источником его может быть квартирантство , хищничество , комменсализм , симбиоз , случайное поселение одного организма на теле другого, заглатывание и другое. Паразиты часто (но не всегда) приносят более или менее существенный вред хозяину и вызывают различные заболевания человека, животных и растений.
Квартирантство, сожительство животных разных видов, основанное на пространственных, а не пищевых связях. При наименее тесном сожительстве — синойкии — квартирант поселяется в жилище хозяина; например, в норах грызунов и др. роющих животных, в гнёздах птиц, муравейниках и ульях пчёл обитает иногда большое число квартирантов. При более тесном сожительстве — эпиойкии — квартиранты поселяются на теле хозяина; например, некоторые питающиеся планктонными организмами усоногие рачки, прикрепляясь к акулам и китам, используют их как средство передвижения. Дальнейшее развитие квартирантства — энтойкия, или квартирантство внутри тела хозяина, при отсутствии пищевых отношений с ним ; примеры: мелкие рыбки Fierasfer, обитающие в клоаке одной из голотурий и периодически выходящие наружу для питания рачками; нематоды, живущие в кишечнике лошади и питающиеся находящимися там инфузориями. Хищничество, форма взаимоотношений между организмами разных видов, из которых один (хищник) поедает другого (жертву, добычу), обычно предварительно убив его. В зависимости от характера взаимоотношений животных, которым свойствен комменсализм , их делят на три группы. 1) Комменсал ограничивается использованием пищи организма другого вида; например, в извивах раковины, занятой раком-отшельником, обитает кольчатый червь из рода Nereis, поедающий остатки пищи рака. 2) Комменсал прикрепляется (временно или постоянно) к организму другого вида, который в этом случае называется хозяином; например, рыба-прилипало своим спинным плавником, превращенным в присоску, прикрепляется к коже акул и др. крупных рыб, пользуясь ими для передвижения; морские гидроиды, поселяющиеся на коже рыб и питающиеся их экскрементами; некоторые ракообразные (например, морские жёлуди), живущие на коже китов и на раковинах моллюсков. 3) Комменсал поселяется во внутренних органах хозяина; например, инфузории опалины, живущие в задней кишке лягушек; некоторые жгутиконосцы, обитающие в кишечнике млекопитающих.
Из комменсализма, характеризующегося поселением комменсала в органах хозяина, мог возникнуть паразитизм. Из комменсализма мог выработаться и симбиоз . Однако комменсализм не является обязательной переходной ступенью к паразитизму или симбиозу; эти формы взаимоотношений организмов могли возникнуть в процессе исторического развития организмов и независимо от комменсализма.
Всё живое в мире находится в тех или иных отношениях друг с другом – от симбиоза и до паразитизма. Многоклеточный организм с этой точки зрения тоже можно рассматривать как вид симбиоза. Попробуем разобраться, как могла возникнуть многоклеточность.
Амеба - простейшее одноклеточное, относящаяся к типу Protozoa. Клетка у амебы выполняет все жизненно необходимые для нее функции. На рисунке показано, как она охотится. Она выпячивает ложноножки, которые охватывают нечто съедобное для амебы.
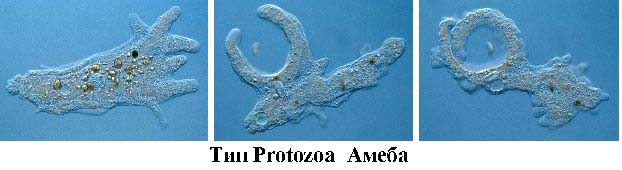
Что должна сделать амеба, чтобы съесть свою жертву? Амеба, в принципе, должна выпустить ферменты (она умеет это делать), которые расщепят жертву на кирпичики. Но проблема состоит в том, что амеба и ее жертва малы по сравнению с той водной средой, в которой все это дело происходит. И если амеба начнет выделять ферменты для того, чтобы растворить часть бактерий в пруду, то она потратит на это энергии больше, чем потом получит от поглощения этой жертвы. Эту проблему амеба решает одним-единственным образом. Она сужает пространство, в котором происходит растворение, и делает его настолько маленьким, что процесс становится энергетически выгодным. Она подползает к своей жертве, обволакивает своим телом и обволакивает ее чем-то вроде пищеварительной вакуоли. Теперь уже становится выгодным выпустить сюда пищеварительные ферменты, все растворить, все нужное взять, а ненужное - выбросить. Эту амебу может съесть теперь только более крупная амеба, которая способна эту, предыдущую амебу, взять в большую пищеварительную вакуоль.
Размер клетки по физическим причинам ограничен весьма небольшими пределами. Так что таким образом – увеличивая размеры клетки – совершенствовать защиту и нападение можно лишь до определенной степени. Даже инфузория, очень крупный представитель простейших имеет врагов. И еще из-за своих размеров она вынуждена иметь дополнительное ядро, чтобы ее клетка нормально функционировала. Вообще говоря, это путь, по которому нельзя идти бесконечно долго. У инфузории, кстати говоря, также есть клеточный рот, и вообще, они с амебой решают одну и ту же задачу – сузить объем среды, в котором происходит пищеварение. Как можно бороться с такими хищниками? Вообще говоря, одним-единственным способом. Если более мелкие амебы (жертвы) образуют комочек из многих клеток, то большая амеба их съесть не сможет.
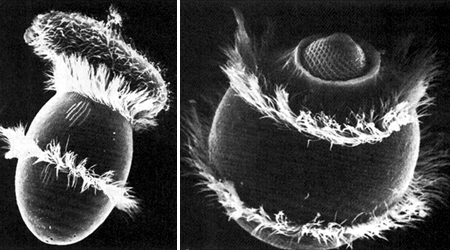
Хищная инфузория дидиниум поедает инфузорию-туфельку
Примером колонии отдельных одноклеточных организмов может служить вольвокс. В принципе, клетка не погибает, если отделить ее от колонии, но при размножении довольно быстро образуется колониальная форма. Колония способна размножаться, то есть внутри колонии образуются более мелкие колонии. До 10 000 одноклеточных может входить в такие колонии. Действия между ними согласуются через контакты между отдельными клетками, то есть все клетки машут жгутиками согласованно, чтобы колония могла перемещаться не беспорядочно.

Но те, кто находится в центре комочка, находятся в плохом положении – они не имеют доступа к еде, и в итоге они гибнут. Таким образом, мы получаем следующую систему – полый шарик, все клетки которого имеют доступ к внешней среде, и в то же время их не может съесть более крупная амеба.
И мы имеем пример такого симбиоза колонии клеток. Именно так устроено примитивное животное губка. Хотя с виду губки похожи на растения, к перемещению не способны, они относятся к животным, потому что фотосинтезом не занимаются и имеют животный тип питания. Стенка у губки имеет трехслойное строение: покровные (толстые наружные) клетки, жгутиковые клетки (хоаноциты, внутренние клетки). Пространство между наружным и внутренним слоями клеток заполнено слизистым веществом, в котором находятся клетки амебоциты (похожи на амебу). В отличие от кишечнополостных, во внутренней полости у губок не происходит пищеварение. Эта полость служит только для протока воды. Жгутики машут таким образом, что вода протекает через поры в теле губки и выходит через устье губки. С водой поступают частицы, которые хоаноциты захватывают и поглощают. Покровные клетки и амебоциты питаются частью съеденной хоаноцитами пищи, которую те им передают. С одной стороны, мы видим дифференцированные клетки, но с другой стороны, они дифференцированы не насовсем: хоаноцит может превратиться в амебоцит, потом перебраться на другую сторону и стать покровной клеткой. Таким образом, нельзя сказать, что губки имеют ткани, как у высших животных: хотя у них и имеются слои дифференцируемых клеток, последние специализированы временно. Еще раз подчеркнем, что тут уже имеется разделение функций между клетками, клетки не одинаковые, но это разделение временно.

Если, говоря о вольвоксе, мы по-прежнему имели дело с представителем Простейших, но уже колониальным, а, рассматривая губки, имели какой-то переходный вариант, то вот отсюда начинает появляться следующая, очень важная конструктивная идея, которая среди ныне живущих организмов реализована у Типа Кишечнополостных. Это гидры, кораллы и полипы. Кишечнополостные реализуют следующую стратегию. Представьте себе мячик, из которого выпустили воздух. Этот мячик можно легко вогнуть в себя. Тогда мы имеем объект такой формы, что все клетки по-прежнему имеют доступ к внешней среде и могут существовать, но при этом мы получаем одно очень большое конструктивное преимущество. Внутри организма образуется полость, которую можно сузить и закрыть, и в ней уже вести пищеварение. В частности, большую амебу можно сюда поместить, закрыть полость и спокойно ее переваривать. Этот тип организации характерен для Кишечнополостных. Если это кишечнополостное будет неподвижно сидеть на дне, то получится полип, если будет плавать по поверхности моря, то получится медуза. Суть от этого не изменится. А изменятся некоторые обязательства, которые животные должны на себя взять. До этого шла речь об организмах, у которых клетки жили просто некоторым случайным собранием и никакие функции друг с другом не делили. Теперь же мы должны разделить обязанности, поскольку реальный доступ к пище имеют внутренние клетки, относящиеся к внутреннему слою, так называемой энтодерме. Но внешние клетки выполняют очень важную функцию. Во-первых, среди них развиваются так называемые чувствительные клетки – нервные клетки. Ведь мы же должны знать, когда нужно сжать эту полость (когда внутрь попала амеба, иначе сжимать ее бессмысленно). С другой стороны, должны быть клетки, которые способны сокращаться, которые могут принять команду «сжать». И получается, что внутренние клетки должны кормить внешние. Таким образом, здесь имеет место то, чего не происходит у таких организмов как губки, - дифференцирование клеток, и разделение между ними функций. Эти самые кишечнополостные животные имеют радиальную симметрию, потому что для них важна только ось «верх-низ» и совершенно не важны стороны света.
 |
 |
 |
|---|
Каким образом клетки, содержащие одинаковый геном, могут иметь разную форму и выполняют разные функции? Для этого должны синтезироваться разные белки, которые идут и на строительство клеток, и на ферментативные функции. Гены во всех клетках одинаковые, за исключением половых клеток. То есть, гены во всех клетках одинаковые, но при этом клетки имеют разную форму и разные функции. Это объясняется тем, что в каждой клетке работают не все гены, а только те, которые нужны в данный момент. Гены могут включаться и выключаться, то есть, как говорят, активироваться либо быть репрессированными (выключенными).
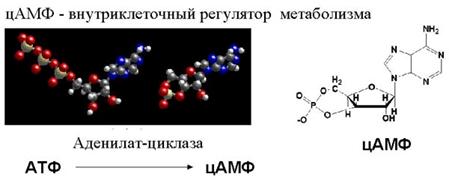
Интересно, что циклический АМФ используется не только для регуляции внутриклеточных процессов, но и для межклеточной коммуникации при формировании многоклеточности. Расскажем об этом на примере уникального организма – амебы, которая называется Dictyostelium discoideum . Это одноклеточная амеба, которая живет в почве и питается бактериями. Иногда её относят к грибам.
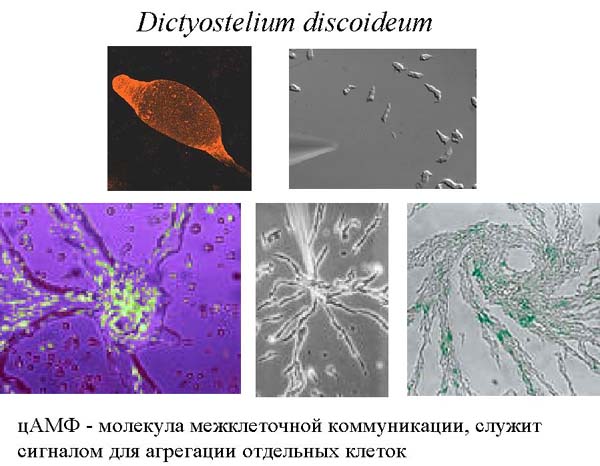
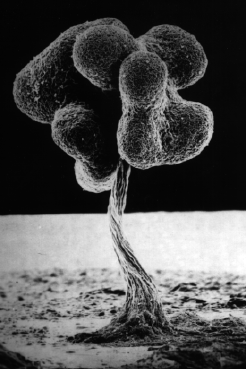
Когда все хорошо, амебы диктиостелиума ползают по своему месту обитания, питаются и делятся время от времени. Но если они голодны, долго не попадалось хорошей еды и их энергозапасы начинают истощаться, они выпускают во внешнюю среду цАМФ, соседние клетки этот сигнал воспринимают. Если клетки сыты, то они на него не реагируют, если же они голодны, то они начинают сползаться в кучу. Вначале они собираются небольшими группами, выпускают циклический АМФ, его концентрация становится больше, поэтому одиночные клетки начинают к ним подползать, образуя агрегат клеток. В итоге к самой большой кучке сползаются остальные группы клеток, и они формируют единый многоклеточный организм, который называется псевдоплазмодий. Миллионы таких клеток собираются вместе и образуют огромную клеточную массу, сильно смахивающую на многоклеточный организм. Он способен ползать, и в отличие от амебы, способен перемещаться на заметные расстояния. Эта масса передвигается и реагирует на свет и химические вещества, словно единое животное. Он с довольно заметной скоростью уползает из плохого места. Если ему удается переползти туда, где есть еда, то он опять распадается на отдельные клетки, которые, как приличные одноклеточные амебы, начинает питаться. Если же он ползет – ползет, а хорошая жизнь все не наступает, то он останавливается, примерно 20% клеток ползут вверх, образуя жесткий стебелек, и затем отмирают (то есть, приносят себя в жертву всем остальным). По стебельку остальные клетки переползают на самую верхушку, образуют плодовое тело, в котором созревают споры. В конечном итоге слизевик предстает в виде плодоносящего тела, во многом похожего на спорангий какого-либо гриба. У него имеется высокая ножка с защитной внешней оболочкой, а сверху располагается мешочек со спорами. Они разбрасываются на некоторое расстояние вокруг, пережидают неблагоприятный период. Когда наступает хорошее время, они прорастают в амебы, и вся история начинается заново. Этот процесс проиллюстрирован на картинке ниже.
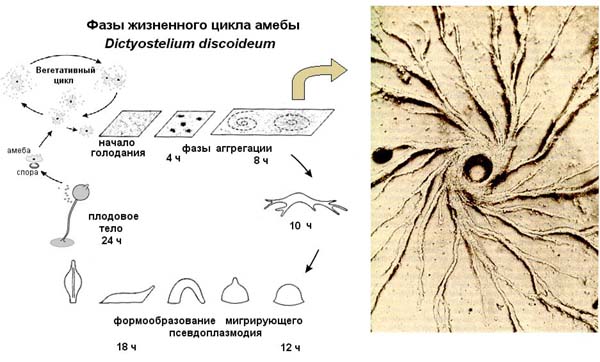
Эта амеба не является ни одноклеточным, ни многоклеточным организмом. В ее случае мы сталкиваемся с тем, что то, что можно наблюдать в живой природе, сложнее, чем придуманная людьми система классификации. Dictyostelium часть своей жизни одноклеточный, другую часть – многоклеточный, и он способен переходить из одной формы в другую, то есть, так просто его не классифицируешь. Он на стадии псевдоплазмодия имеет дифференцированные клетки. Его передний конец обладает хеморецепцией, он лучше все чувствует, чем задний конец псевдоплазмодия. И образование плодового тела – это процесс уже ярко выраженной дифференциации, при которой 20 % клеток образует стебелек и погибают. Размер генома Dictyostelium 'a 3.5x107 bp. Мы можем назвать Dictyostelium факультативно многоклеточным.
Итак, благодаря тому, что не все гены работают в клетке одновременно, клетка может менять программу своей активности, образовывать разные ферменты и иметь разную форму. Это происходит как у одноклеточных, так и у многоклеточных. У многоклеточных регуляция еще более сложная, так как помимо внутриклеточных процессов нужно регулировать еще межклеточные взаимодействия. Но возникает вопрос, как вообще могли образоваться многоклеточные. Как исходно одноклеточные организмы превратились в многоклеточные? У практически всех одноклеточных известны мутанты, не расходящиеся при делении клетки. То есть нормальное деление клетки происходит, все у нее делится: ядра делятся, митохондрии делятся, хлоропласты расходятся по разным дочерним клеткам, но последний этап, когда клетки должны отцепиться друг от друга, у них не происходит, они остаются сцепленными. У некоторых видов эти клетки-мутанты живут хуже, чем нормальные одноклеточные формы, потому что у них нет системы регуляции взаимодействия клеток. Но некоторые виды, видимо, приспособились к этому, эти клетки начинают взаимодействовать друг с другом, у них есть программа согласования своих действий. Нерасхождение при делении встречается не только на уровне одноклеточных, но и на уровне многоклеточных, (например, сиамские близнецы). Не всегда это является уродством, иногда это вариант нормы.
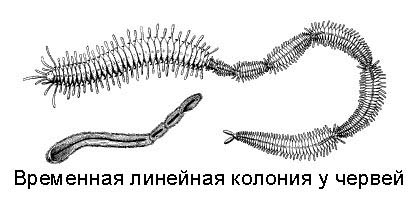
На рисунке вы видите червей, которые после деления не разошлись. Все вместе они образуют то, что называется временной линейной колонией. Если их друг от друга отделить, они будут дальше вполне комфортно жить. Такая колония может потом сама разделиться. Но происходит это не сразу, какое-то время черви живут все вместе.
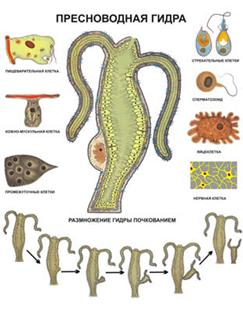
Существуют колонии так называемых гидроидных полипов. Напомним, что гидра относится к типу Кишечнополостных, подцарство Metazoa. Тело гидры имеет вид удлиненного мешочка. Его внутренность — кишечная полость — сообщается с внешней средой через ротовое отверстие, окруженное несколькими щупальцами. Стенки мешочка состоят из двух слоев клеток: внутреннего ( энтодерма ) и внешнего ( эктодерма ). И в эктодерме, и в энтодерме много мышечных клеток, содержащих волоконца, которые могут сокращаться, приводя тело гидры в движение. Кроме того, в эктодерме есть и нервные клетки, причем клетки, расположенные ближе всего к поверхности, — это рецепторы, а клетки, заложенные глубже, среди мышц, — эффекторы. Если к гидре прикоснуться иглой, она сжимается в комочек. Это простой рефлекс, вызванный передачей возбуждения от рецепторов к эффекторам. Но гидра способна и к гораздо более сложному поведению. Захватив добычу, она подтягивает ее щупальцами к ротовому отверстию и заглатывает.
Такое строение имеют и остальные представители кишечнополостных.
Многие виды кишечнополостных образуют колонии. Ниже представлена колония животных – гидроидных полипов, которая с виду похожа на растение. Каждый "листочек" такого "растения" имеет такое же строение, как гидра.

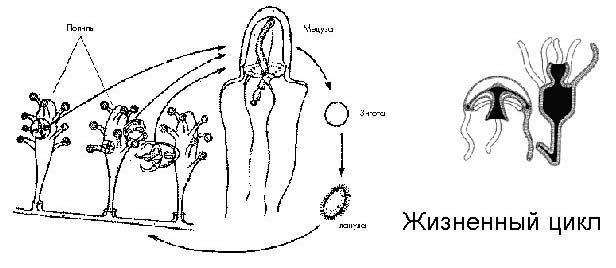
Жизненный цикл у некоторых из гидроидных полипов проходит следующим образом: полипы растут вместе, они объединены общим проводящим каналом, время от времени некоторые из них отрываются, превращаются в медуз (принципиальное строение гидры и медузы одинаковое), которые уплывают и активно размножаются, проходят определенные стадии, образуют половые клетки, которые, сливаясь, дают зиготу, из которой вырастает новый полип. И снова весь цикл повторяется.
Известные нам красивые кораллы – это скелет коралловых полипов. Полипы – похожие на гидру существа, у которых есть общий проток воды по общим объединяющим их каналам, они строят для себя известковый скелет. У некоторых этот скелет включает в себя соединения железа, которые окрашивают его в красивый красный цвет. Коралловые полипы живут в море. На следующем рисунке представлены разные формы коралловых полипов.

На следующем рисунке слева представлен мадрепоровый коралл. У него отдельные полипы уже неразличимы, они все сливаются в единое целое, от них остаются только глотки с щупальцами, причем эти щупальца обобществлены, они заглатывают кусочки пищи и направляют ее в общую полость. Справа виден родственник гидры и полипа – мшанка кристателла. У нее тоже отдельные особи сливаются своими основаниями. Каждая особь называется не полипом, а зооидом, то есть это как бы и не отдельное животное, и не орган. Они способны втягиваться в трубку у основания общего тела. Объединенная часть их образует подошву, и, в отличие от кораллов, мшанка способна передвигаться со скоростью 15 мм в сутки.

Другой более отдаленный родственник гидры - физалия - относится к подклассу сифонофор. Это животное называют португальский военный кораблик. У физалии есть сверху парус, который может иметь разную окраску, от голубого до пурпурного. Она довольно ядовитая, у нее есть стрекательные клетки, которые способны сильно отравить даже человека, не говоря уже о мелких животных, которыми она питается. На рисунке представлена схема строения сифонофор. Тело сифонофоры устроено из группы зооидов, разным образом модифицированных.
Самый верхний образует плавательный пузырь, внутри него находятся клетки, выделяющие газ, если физалии надо подняться на поверхность. Под плавательным пузырем находятся плавательные колокола – это как бы отдельные индивиды, но здесь они уже превращаются в органы. Ниже находятся индивиды, которые используются как половые органы, они не способны питаться, поэтому другие зооиды – гастрозоиды, их кормят, но не только их, но и весть «организм». Есть защитные зооиды, имеющие стрекательные клетки.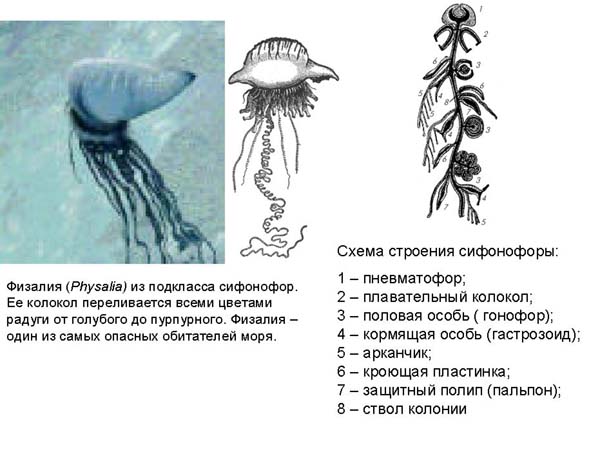
Таким образом, если у колонии коралловых полипов каждый полип равнозначен, то здесь колония может превращаться в достаточно сложноустроенный организм, с дифференцированными полипами, каждый из которых превращается в орган. Таким образом, этот организм нельзя однозначно проклассифицировать – является ли он колоний организмов или отдельным организмом.
Аналогичную ситуацию мы можем наблюдать, изучая социальных насекомых – муравьев, пчел, термитов.
Можно ли назвать колонию муравьёв, пчел или термитов организмом или это всё-таки совершенно отдельные независимые особи? Если это независмые особи, то они должны быть способны к самостоятельному существованию вне колонии и иметь полный комплект функций своего организма. Так ли это?
У пчел (так же как и у муравьев и термитов) обычно имеется единственная плодоносящая матка. Остальные пчелы – рабочие самки, они неплодоносящие и по сравнению с маткой мелкие. Они кормят матку, которая ничем другим не занимается, кроме как откладывает яйца, из которых появляются рабочие пчелы или трутни.

Они строят огромные термитники. Справа сверху показаны: 2 рабочие особи, солдат и матка. Пока она функционирует, другие особи не могут развиться в матку. Если ее убрать, то одна из рабочих особей превращается в царицу – плодовитую матку. Это регулируется (так же как и у пчел и муравьев) через выделение ферамонов и др. веществ матки. То есть матка выделяет вещества, которые ингибируют (запрещают) трансформацию других самок в матку. Когда матка умирает, все самки начинают выделять эти вещества, и та самка, которая опережает других, тормозит их, становясь маткой. Это означает, что у самок не утеряна способность плодоносить, но она репрессирована.
У термитов, если есть вторая плодоносящая самка, то она улетает в другое место и образует новое сообщество. Одна оплодотворенная матка может создать целое гнездо. Она хранит спермии в определенном резервуаре, которые расходует для оплодотворения яиц очень экономно. После того, как она отложит яйца, она за ними начинает ухаживать, как только появляются личинки, матка их кормит частью яиц, и после этого, личинки превращаются в рабочих особей. Они начинают помогать матке кормить новых личинок вначале яйцами, затем добычей. Часть из рабочих особей ходят за добычей, а часть строят термитник. Таким образом от одной плодоносящей матки может получиться новое гнездо с огромным количеством особей.
У пчел матка может откладывать как оплодотворенные, так и неоплодотворенные яйца. Из первых образуются рабочие пчелы-самки, из вторых – трутни. Трутни могут вылетать в другие ульи и оплодотворять чужую матку, таким образом осуществляя перекрестное оплодотворение. Хотя можно использовать трутней из своего улья, потому что при этом все равно происходит перекомбинация генетического материала.
Здесь следует подчеркнуть, что вегетативное деление клетки вполне обеспечивает увеличение числа клеток. Половой процесс нужен для перекомбинации генетического материала. Есть виды, например среди одноклеточных, для которых описано самооплодотворение. Когда заканчивается обычное деление, проходит половой процесс. Клетка делится, образуя половые клетки, которые потом сами с собой сливаются, то есть потомки одной клетки сливаются друг с другом и образуют зиготу. В данном случае деление не приводит к увеличению числа потомков, но у них перемешивается генетический материал.
Какую можно провести аналогию между термитами и, например, сифонофорами? Мы здесь видим отдельных индивидов, вроде бы независимых друг от друга, но они образуют колонию, хотя в отличие от сифонофор, отдельные части этой колонии способны самостоятельно передвигаться. Но ясно, что колония – это «организм» с дифференцированными органами. Этими органами являются отдельные термиты. Есть репродуктивный орган – матка, орган защиты – солдаты, есть органы питания и ухода за личинками – рабочие самки, есть система коммуникаций (химическая и поведенческая формы) между органами. Как же социальные насекомые между собой общаются?
В свое время было сенсацией расшифровка языка пчел.
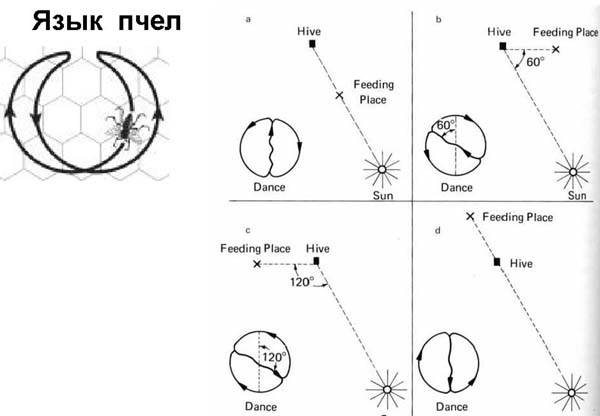
Когда пчела нашла нечто съедобное, она прилетает обратно в свой улей и сообщает остальным, что и где она нашла. Что она нашла, понятно по тому, что прилипло к ее телу. А где она это нашла – сообщает на языке танца. Во время танца пчела движется по вертикальной поверхности. Она совершает движение, схематически отражающее направление и расстояние до места, куда следует лететь за едой.
Солнце в этой схеме по умолчанию находится сверху. Например, если место кормежки расположено прямо по Солнцу, то пчела ползет вверх и делает такую «восьмерку», как показано на рисунке. Таким образом направление она указала. Если место для питания под углом 60 0 , то пчела поворачивает восьмерку на 60 0 (рисунок b). Аналогично остальные ситуации. А количеством виляний брюшком она указывает на расстояние. У разных пчел разные диалекты, поэтому у них одно число виляний брюшком показывает разное расстояние. Также пчела может объяснить не только, как лететь по прямой к месту кормежки, но и как облететь, например, гору. Если еда находится рядом с ульем, то пчела выполняет танец, который показан на рисунке слева вверху.
Как же пчела ориентируется на местности, как запоминает ориентиры? Дело в том, что пчелы видят все в поляризованном свете, у них по другому устроено зрительное восприятие, и есть посвященные этому исследования. Интересно знать, что у пчел врожденное владение языком. При жизни они, конечно, совершенствуют свой танец, добавляют новые, свойственные только этому улью элементы танца, но в принципе они изначально могут показать путь к месту, где есть еда. Если взять пчел, говорящих на разных диалектах, то оказывается, они не могут понять друг друга. Возникает вопрос: применяют ли пчелы еще в каких-нибудь ситуациях свой «язык»? Нет. Для остальных ситуаций у пчел хорошо развита система коммуникаций, регулирующая отношения индивида внутри роя, для того, чтобы они делали то, что надо. Но наиболее загадочным и интригующим для ученых была их способность рассказать о том, куда лететь. За работу над расшифровкой языка пчел Карл фон Фриш получил Нобелевскую премию (1973 г.).
Хочется отметить один факт: что у нас, что у пчел общая архитектура нейронов задана наследственно, а не обучением, индивидуальный опыт добавляет то, как они друг с другом соединятся на конечных стадиях. Мы отличаемся от пчел тем, что у нас гораздо большая часть нервных связей наследственно не задана, и на них ложится индивидуальный опыт.
Существуют две причины, почему отдельные независимые индивиды (будь это клетки или гидроидные полипы, или приматы) объединяются в колонию или многоклеточный организм. Это наиболее эффективное использование ресурсов и защита. На примере волков объясним первую причину. Летом волки живут небольшими группами, семьями. Они занимаются тем, что ловят разную живность, например мышей, которых летом много, и вполне успешно способны прокормиться. Но зимой мыши и другие мелкие грызуны прячутся под снегом, и приходится ловить крупных животных – лосей или оленей, например. Тогда волки сбиваются в стаи. Стаи могут осуществлять очень сложные виды загонной охоты, некоторые загоняют, некоторые сидят в засаде. Во время охоты волки обмениваются между собой вокальными сигналами, координируя таким образом свои действия. То есть здесь мы сталкиваемся с ситуацией, аналогичной поведению Dictyostelium 'а, который часть времени проводит как одноклеточное, а часть времени – как коллектив одноклеточных, объединяющихся в единый организм
Очень интересен образ жизни приматов. Шимпанзе живут отдельными группами, которые включают в себя несколько десятков особей, они ведут образ жизни охотников-собирателей, питаются они растениями, мелкими животными, объединяются в группы для охоты, несколько молодых самцов способны поймать мелкую добычу и съесть, также они едят муравьев, термитов, и используют орудия для того, чтобы их ловить. Например, для того, чтобы достать термитов, они разжевывают веточку, опускают в термитник, там на нее налипают термиты, и шимпанзе вынимает веточку и снимает их с нее губами. Для того, чтобы достать воду, они способны использовать губку из травы. Они могут разбивать орехи двумя камнями, один из которых – прототип молота, второй – наковальни. В группе существует достаточно строгая иерархия: есть лидирующий самец. Лидер выбирается не по силе и агрессивности, как раньше думали, а по способности к образованию максимального числа дружеских связей и способности к организации своих соплеменников на кооперативные действия.

У человека сохранились от наших родственников приматов некоторые формы поведения.
Первые эукариоты появились около 2 миллиардов лет назад. Многоклеточные появились, видимо, более 600 миллионов лет назад, данные об этом появились тоже благодаря ископаемым породам. Что же касается млекопитающих, то они появились в эволюционных масштабах недавно, всего 200 млн лет назад.
Ближайшим родственником человека, имеющий общего с нами предка, считается шимпанзе. Шимпанзе и человек имели общего предка 5 миллионов лет назад, примерно в это время ветви, ведущие к человеку и шимпанзе, разделились. Около 2 миллионов лет назад у человека появились первые примитивные орудия труда. Предком человека современного типа был вид Homo erectus (человек прямоходящий). Человек современного анатомического типа появился примерно 100-150 тысяч лет назад. Представления о происхождении человека, его родстве с представителями живого мира были получены при сравнении строения тела человека и других животных, данных сравнительной эмбриологии, исследования костных останков вымерших видов – данных археологии и палеонтологии, а за последние десятилетия также и при сравнении ДНК разных организмов. На основе этих данных восстановлен общих ход развития жизни на Земле. Однако ответить на вопрос, почему жизнь появилась; почему она усложнялась; что способствовало ее усложнению; каким образом из аминокислот, нуклеотидов и другого материала получилась такая сложная функционирующая система, почему очень просто устроенные организмы в ходе эволюции все более и более усложняли свою структуру трудно. Эти вопросы обсуждаются в теории эволюции. Дарвин был не первый, кто выдвинул идею эволюции, но он сумел обобщить и преподнести те идеи, которые существовали до него, так, чтобы они были восприняты обществом, и сумел развить их в своей теории. Причем его работа попала в круг всеобщего внимания не только среди ученых, но и среди широкой публики. В то время, когда основной темой для диспутов в ученых кругах была теория естественного отбора (то есть, эволюционируют ли виды под действием естественного отбора или по другой причине), общественность обсуждала вопрос о том, от кого же произошел человек: от Бога или от обезьяны.
Со времен теории Дарвина было проведено много новых исследований, возникли новые теории эволюции. Основатель учения, называемого антропософией, Штайнер, считал, что эволюция идет не от простого к сложному, а наоборот. Объединение идей генетики и теории эволюции привели к появлению так называемой синтетической теории эволюции, которая ответила на многие вопросы. Но, тем не менее, есть вопросы, на которые и эта теория не может ответить. Есть направление, называемое креационизмом, последователи которого утверждаются, что все виды созданы Богом. Они неизменны, не могут совершенствоваться. Существование палеонтологической летописи, то есть появление и исчезновение видов в истории Земли, с точки зрения креационистов следует объяснить тем, что Бог поместил эти виды на Землю, а потом изъял их. Креационисты проводят достаточно интересные расчеты, связанные, например, с тем, какие и сколько видов животных имел возможность Ной собрать в своем ковчеге, каких размеров должен быть этот ковчег. Рыб и растений, по их теории не надо было брать, так как они могли выжить в воде сами. С их точки зрения, Ной мог взять молодых животных, которые были более жизнеспособны и занимали меньше места, таким образом, они высчитали определенный минимум видов животных, который обеспечил разнообразие жизни на Земле сегодня. Креационисты находят слабые места в дарвиновской теории эволюции, задают вопросы, ответы на которые эволюционистам приходится искать.
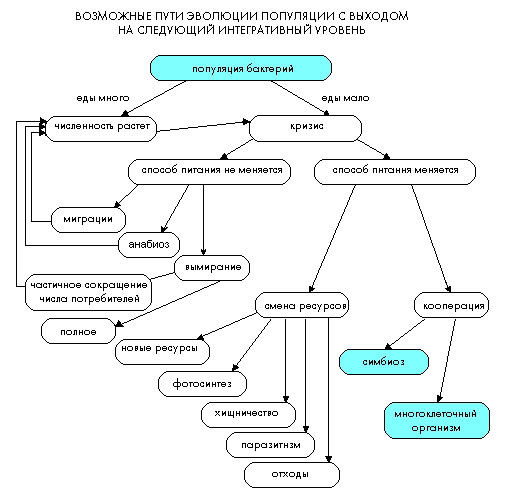
Для того, чтобы представить, как могла проходить эволюция и какими могут быть ее причины, построим следующую модель, и попытаемся понять, почему живые организмы усложнялись. Пусть, для простоты модели, у нас есть один вид бактерий, который питается одним видом пищи. Будем наблюдать за тем, как ведут себя эти бактерии в разных ситуациях. Ясно, что пока есть благоприятные условия и много еды, бактерии усиленно размножаются. Рассмотрим период жизни, когда бактерии съели всю пищу, еды больше не осталось. Что может происходить с бактериями в сложившейся ситуации? Из этой ситуации есть 2 выхода, связанные с тем, меняется или нет способ питания:
1. Если способ питания не меняется, то у бактерий есть 3 выхода:
1.1. Мигрировать на территорию, где есть много еды, это опять приведет к увеличению численности и таким образом круг замкнется (для того, чтобы мигрировать, необходимо обладать достаточной подвижностью, то есть возникают приспособления для движения).
1.2. Второй выход – это вымереть:
1.2.1. если вымирание частичное, то после кризиса с едой, численность бактерий возрастет и круг опять замкнется (такие циклы происходят очень часто (модель волка и зайца), когда численность бактерий превышает количество ресурсов, из-за этого происходит частичное вымирание вида до момента, когда еды опять начинает хватать всем, после этого опять возрастает численность организмов и т.д.);
1.2.2. вымирание полное ведет к полному исчезновению вида.
1.3. анабиоз («впасть в спячку» до лучших времен).
2. Если способ питания меняется, то у бактерий есть 2 пути развития:
2.1. сменить ресурсы путем:
2.1.1 выработки органических веществ самим (фотосинтез);
2.1.2. хищничества (поедание себе подобных);
2.1.3. паразитизма («откусывание по кусочку» от живого организма);
2.1.4. использовать отходы (пример: раньше атмосфера Земли была восстановительная, и для живых организмов кислород был сильным ядом, но постепенно, по мере накопления кислорода в атмосфере, выживали только те, кто освоил аэробный тип дыхания);
2.2. можно кооперироваться (для этого надо уметь общаться, у клеток это происходит на уровне электрических сигналов, проходящих по мембранам клеток (прообраз нервного импульса), и химических сигналов (прообраз гормональной системы).
2.2.1. примером кооперации является симбиоз (один из важнейших факторов эволюции);
2.2.2. при кооперации одинаковых клеток возникает многоклеточный организм - в нашем случае многоклеточные бактерии. Такие бактерии действительно существуют.
И симбиоз, и интеграция клеток одного вида приводит к повышению сложности, переходу на новый интегративный уровень развития. При этом вырабатывается система связи между элементами предыдущего уровня, то есть, если ранее говорилось об отдельных бактериях, совокупности скооперированных бактерий, то сейчас уже речь идет о системе взаимодействующих клеток.
Эукариоты появились в результате симбиоза бактерий. Есть организмы (например, амебы Pelomyxa palustris), которые представляют собой целую команду. Вместо митохондрий в данном организме живут бактерии; вместо жгутиков – другие бактерии (причем, два вида - длинные и короткие), которые, синхронно поворачивая хвостами, обеспечивают этому организму движение. Сейчас есть множество примеров того, что некоторые живые организмы зафиксировали собой как бы промежуточную стадию (тупиковую) эволюции, не переходя на новый интегративный уровень.
Теперь представим, что на "входе" в данной схеме вместо «популяции бактерий» поставлены «многоклеточные организмы». Очевидно, что многоклеточные будут иметь те же принципиальные возможности выхода из кризисной ситуации, а в результате интеграции появятся колонии, стаи и т.п.; другими словами – группа взаимодействующих многоклеточных организмов, которую мы назовем первичным социумом (это не устоявшийся термин), или социальный организм. Если теперь вверху схемы поставить «первичный социум», то, в итоге, получится система социумов, которая, в зависимости от структуры, называется племя или вождество. Этот этап эволюции характерен только для человека. С помощью таких схем, можно проследить переходы организмов на новые интеграционные уровни.
В качестве примера социального организма можно также рассмотреть пчел и муравьев. Но, в данном случае, они представляют собой скорее многоклеточный организм (у которого «ручки» и «ножки» бегают отдельно). Рассмотрим первичный социум, например, стадо обезьян. Стадо наших ближайших родственников, шимпанзе очень похоже по своей структуре на первобытную общину. Первобытная община имела такую же структуру как группы охотников-собирателей в Африке, группы бушменов. Это небольшие объединения (человек 20-40, редко больше), кочующие по своей территории. За каждой общиной закреплена определенная территория; другие племена на ней находиться не могут; размер территории (и общины) определяется следующим фактором: сумеют ли они за год собрать необходимую пищу с территории. Соответственно, чем менее плодородна земля, тем численность общины меньше, и наоборот, чем более плодородна почва, тем численность общины больше, а территория, соответственно, меньше. При объединении первобытных общин возникает более сложная социальная структура – племя. На самом деле существует еще одна структура – вождество. Этот термин узкоспециализированный. От племен вождества отличаются тем, что они более иерархизированы. Племена - это союз общин, и в них, в общем-то, нет вождя, который может кому-то что-то приказать. Во главе вождеств стоит вождь, который всем руководит и который принимает дары от членов вождества и перераспределяет их. В результате объединения племен или вождеств возникает государство; а объединение государств приводит к появлению различных интернациональных союзов и организаций.
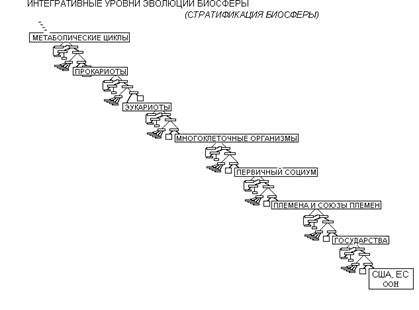
Следует отметить, что симбиоз – это процесс, который будет идти всегда. На данный момент в разных группах он находится на разных фазах развития.
А теперь посмотрим, что будет в начале этой схемы, если «на выходе» будет прокариотическая клетка. Тогда на "входе" будет система метаболических циклов. Если многоклеточный организм – это система общающихся между собой клеток, то прокариотическая клетка – это система общающихся между собой метаболических циклов. Метаболические циклы включают в себя процессы анаболизма (расщепление более сложных веществ на простые) и катаболизма (объединение простых веществ в более сложные). У нас с растениями и бактериями есть некоторые общие процессы, поэтому, в частности, растения и лечат. Все процессы идут согласовано друг с другом, так как продукты одних реакций являются субстратами следующих, и таким образом, скорость всех реакций сопряжена.
Ниже представлен цикл Кребса – основной энергетический цикл клеток. Этот цикл обеспечивает клетку энергией, в его ходе синтезируется АТФ.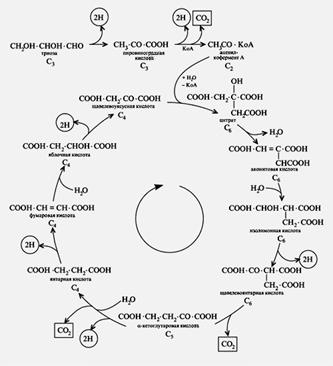
Как могла возникнуть такая сложная система? Есть некоторые ученые, считающие, что жизнь на Землю была занесена из Космоса. Эта концепция называется теорией панспермии. Они считают, что на Земле было слишком мало времени, чтобы возникла такая сложная система, как живая клетка. И в качестве иллюстрации абсурдности подобных предположений проводят следующее сравнение. Дескать, у обезьяны, усаженной за пишущую машинку, больше шансов отпечатать сонет Шекспира, чем у жизни возникнуть на Земле из аминокислот, нуклеотидов и других веществ, из которых состоит клетка. Но у данного сравнения есть слабое место. Рассуждая по аналогии, приведем следующий пример. Шекспир, в отличие от обезьяны, складывал свои сонеты не из отдельных букв. В его распоряжении были готовые сюжеты, которые он заимствовал из других произведений, он пользовался английской грамматикой, знал правила драматургии. То есть он творил не из отдельных элементов, а из готовых блоков.
Концепция блочной эволюции как раз и утверждает, что эволюция происходит не из отдельных молекул, каждый раз создавая что-то новое практически с нуля, а из уже готовых блоков. Как же могла возникнуть жизнь? На самом деле, метаболические циклы возникли тоже не сразу. Можно выделить несколько уровней реакций. И вот самые простые циклы сопрягались с другими, и получались системы биохимических реакций, циклов. Существует интересная теория, описывающая возможный процесс возникновения самого первого такого цикла, ставящая под сомнение необходимость занесения готовых процессов из Космоса. Когда Земля еще только остывала и не было еще никаких бактерий, но Земля вращалась вокруг своей оси, и на нее светило Солнце. Существовали уже ночь и день (возможно, также зима и лето). И «ночью» происходили процессы конденсации, а «днем» - возгонки и конвекции. Возможно, эти процессы, были не чисто физическими, но также и химическими. То есть происходили следующие процессы: на свету, «днем», синтезировались вещества, которые в тени, «ночью», распадались на более простые. И такие циклические реакции могли происходить в масштабах всей планеты. Если процесс днем "успевал" запасти столько энергии, что за ночь не "умирал", то есть преобразуемые в нем вещества не распадались за ночь, на следующий день этот же процесс мог восстановиться. Такие процессы могли представлять собой первые метаболические циклы. Согласно этой концепции жизнь (самовоспроизводящиеся процессы) возникла до появления живых организмов. Это только гипотеза, она не является общепринятым мнением. Существует журнал «Молекулярная эволюция», который в том числе рассматривает вопросы происхождения жизни. Наибольшее внимание в нем уделено происхождению объектов, вопросам о том, какая молекула из какой возникла. Вопросам возникновения процессов посвящена лишь маленькая толика всех исследований. Однако эволюция процессов не менее, а может, и более интересна, чем эволюция объектов. Возможно, не всегда нужно рассматривать какую-то конкретную молекулу, а более полезно разобраться в каких процессах она участвует. Это напоминают историю про курицу и яйцо (что было раньше: курица или яйцо?): понятно, что это процесс, а в отдельности курица и отдельно яйцо – это два разных состояния этого процесса, как бы две промежуточные стадии бесконечного процесса.
Возможно ли сейчас абиотическое появление клетки? Сейчас это невозможно. Если бы существовало какое-то скопление органических веществ, то оно было бы съедено уже существующими клетками. Почему сейчас обезьяна не превращается в человека? Один вариант ответа: она превращается, но мы просто не замечаем; второй вариант: существующие люди ей это сделать просто не дадут, так как занимают пригодные для этого экологические ниши. Как и государства: если, где-нибудь захотят создать государство, как это когда-то было, то уже существующие на этой территории государства не позволят это сделать. Возможно деление уже существующих государств, но появление новых не реально.
А почему же все-таки происходило повышения уровня организации, почему возрастала сложность систем? Бактерии размножаются и передают ДНК из поколения в поколение ни чуть не хуже других организмов. Зачем надо было повышать сложность организации, а не решать задачи в рамках уже имеющейся организации? Что же служило источником усложнения? Рассмотрим следующий пример.
Если закручивать веревку, то через некоторое время на ней начнут появляться витки второго порядка, а потом и третьего. На этом примере видно, что чтобы в системе что-то происходило, необходимо, чтобы в нее поступала энергия. А когда «емкость» системы данного уровня сложности исчерпана, т.е. упругость нашей веревки больше не позволяет образовываться виткам, происходит образование витков второго порядка и т.д. Это – аналогия, иллюстрирующая то, что все жизненные процессы на Земле, в том числе эволюция, с усложнением структуры систем, происходят при поступлении в систему энергии. Есть два источника энергии – солнечная энергия и тепло недр земли. Мы сейчас, вряд ли можем оценить, какой источник был более важным, какой сыграл более важную роль, но понятно, что оба они необходимы для жизни. Если бы энергия не поступала, то жизнь бы прекратилась, и никакой эволюции бы не было. Если бы она поступала медленно, то все эволюционные процессы протекали бы очень медленно. Если бы она поступала очень быстро, то все бы разрушилось. То есть поток энергии должен быть таким, чтобы он был способен поддерживать биологические процессы, не разрушать молекулы. Часть постоянно поступающей энергии рассеивается, часть идет на поддержание существующих структур, эту энергию перерабатывающих, и часть – на образование новых структур из уже существующих, упорядочивание, повышение эффективности их взаимодействия. То есть, в эволюции участвуют вещество, энергия, а в результате их взаимодействия все структурируется. Можно сказать, здесь возникает порядок из хаоса. Назовем этот третий параметр информация. Из-за того, что Солнце светило на Землю, а в Земле происходили процессы, согревающие ее поверхность, потоки вещества структурировались с образованием все более сложных систем. Можно сказать, что в информации, т.е. в структуре потоков веществ, которые составляют жизнь, законсервирована солнечная энергия, поступавшая на Землю.
В большинстве курсов биологии в качестве одного из основных признаков отличия прокариот от эукариот называется наличие у последних двухмембранных органелл (митохондрий и пластид). Эти органеллы, помимо двойной мембраны, имеют еще целый ряд характерных признаков, которые выделяют их среди остальных клеточных мембранных образований. Вопрос их происхождения неразрывно связан с вопросом происхождения эукариот. Ответ на этот вопрос дает теория симбиогенеза.
Идея о том, что некоторые органоиды клетки могут быть симбиотическими организмами, возникла в начале ХХ века в России. Автор ее - хранитель Зоологического кабинета Казанского университета К.С. Мережковский. Этому предшествовало установление Фаминцыным и Барановским симбиотической природы лишайников (1867). То, что лишайники - продукт симбиоза, некоторые ботаники не признавали и через 50 лет! Очень уж непривычно, что такой "знакомый", милый сердцу организм - не "сам по себе", а сращение двух других организмов.
 |
 |
|---|---|
| Поперечный разрез слоевища гетеромерного лишайника. |
1 — уснея цветущая (Usnea florida); 2 — лобария лёгочная (Lobaria pulmonaria). |
 |
 |
 |
|---|
Та же ситуация имела место и с идеями Мережковского. Хлоропласты - не части клетки, а самостоятельные организмы?! Наши клетки напичканы бактериями - митохондриями?! И дышим-то не мы сами, а они?! Эту теорию не признавали тоже 50 лет. Однако потом появились последователи - уже в Америке. Накопились новые данные.
 |
 |
|---|---|
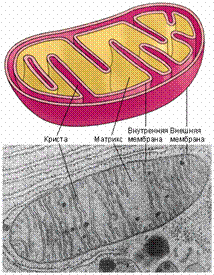
Митохондрии. |
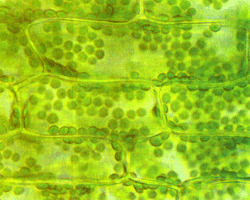
Хлоропласты в растительных клетках. |
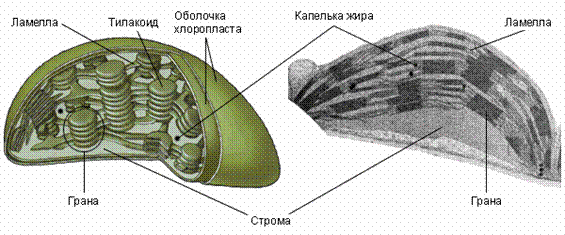
Строение хлоропластов. Хорошо видны содержащие хлорофилл граны, собранные из стопки тилакоидных мембран. Справа – электронная фотография.
Пластиды – органеллы, свойственные только растительным клеткам. Они окружены двойной мембраной. Пластиды делятся на хлоропласты, осуществляющие фотосинтез, хромопласты, окрашивающие отдельные части растений в красные, оранжевые и жёлтые тона, и лейкопласты, приспособленные для хранения питательных веществ: белков ( протеинопласты ), жиров ( липидопласты ) и крахмала ( амилопласты ).
Пластиды обладают относительной автономией. Так же, как и митохондрии, образующиеся из предшествующих митохондрий, они рождаются только из родительских пластид. Причина заключается в том, что эти органеллы содержат небольшое количество собственной ДНК. Подобная внехромосомная наследственность не подчиняется менделевским законам. Анализ мутаций показывает, что ДНК органелл отвечает лишь за малую часть наследственной информации. По-видимому, пластиды также произошли от симбиотических прокариот, поселившихся в клетках организма-хозяина миллиарды лет назад.
Точку зрения на митохондрии и хлоропласты, как на приобретенные клеткой симбиотические бактерии подтверждает ряд особенностей строения и физиологии этих органелл:
1. У них есть все признаки "элементарной клетки":
- полностью замкнутая мембрана;
- генетический материал - ДНК;
- свой аппарат синтеза белка - рибосомы и др.;
- размножаются делением (причем делятся иногда независимо от деления клетки).
2. У них есть признаки сходства с бактериями:
- ДНК обычно кольцевая, не связана с гистонами;
- рибосомы прокариотические - 70S-типа и мельче. Нет 5,8S-рРНК, характерной для эукариот;
- рибосомы чувствительны к тем же антибиотикам, что и бактериальные.
Однако у хлоропластов и митохондрий нет клеточной стенки, характерной для предполагаемых предковых групп. Но ее нет или почти нет и у многих современных эндосимбионтов. Видимо, она теряется для облегчения обмена между симбионтом и хозяином. Кроме того, у водоросли Cyanophora paradoxa найдена "промежуточные форма" - так называемые цианеллы. Эти органоиды (симбионты?) имеют редуцированную клеточную стенку, почему их и считают цианобактериями. В то же время они имеют размер генома, в 10 раз меньший, чем бактериальный (что характерно для хлоропластов) и не размножаются вне клетки-хозяина. Примечательно, что Cyanophora - жгутиковая водоросль, а ее цианеллы больше всего напоминают хлоропласты красных водорослей, которые жгутиков всегда лишены.
А вот затруднение из трудных. Многие белки митохондрий и хлоропластов кодируются ядерными генами, синтезируются на рибосомах цитоплазмы и только потом доставляются аж сквозь две мембраны в органеллу! Как так могло получиться? Единственное объяснение в рамках симбиогенеза - часть генов органелл переместилась в ядро. Еще двадцать лет назад казалось, что это - чистый бред. Потом накопились данные о мобильных генетических элементах (одним из этапов знакомства с ними научной общественности в нашей стране стала книга Р.Б. Хесина "Непостоянство генома"). Гены меняют места в хромосомах, вирусы встраиваются в геномы бактерий и эукариот и т.п. Процесс перемещения генов в ядро стал казаться более вероятным, но ничем не был доказан. Однако позднее появились и свидетельства того, что такой процесс, видимо, действительно имел место. Помогли белки из нескольких аминокислотных цепей. Один из таких белков митохондрий - протонная АТФ-синтетаза - состоит из 8 субъединиц (пептидных цепей). И вот оказалось, что у дрожжей 4 закодированы в митохондриях, а 4 - в ядре. Это уже само по себе подозрительно! А у человека все 8 цепей закодированы в ядре. Значит, в ходе эволюции эукариот от общих предков дрожжей и человека гены переместились в ядро - значит, это в принципе возможно.
Среди современных прокариот паразит растений Agrobacterium tumifaciens, проникая в клетки, встраивает свои плазмиды в геном хозяина, вызывая рак - трансформацию клеток. Этот пример доказывает возможность переноса генетического материала из прокариотической клетки в ядро эукариота-хозяина.
С помощью теории симбиогенеза были предсказаны и/или объяснены многие признаки митохондрий и хлоропластов. Некоторые из предсказаний, которые рассмотрены ниже, сейчас - скорее доказательства: они подтвердились.
Прежде всего, теория симбиогенеза объясняет наличие двойной мембраны и ее свойства. Приобретение двойной мембраны - результат фагоцитоза; наружная мембрана - бывшая мембрана пищеварительной вакуоли и, таким образом, принадлежит хозяину, а не эндосимбионту. Хотя сейчас эта мембрана воспроизводится вместе с органоидом, как ни странно, по липидному составу она больше похожа на мембрану эндоплазматической сети клетки, чем на внутреннюю мембрану самого органоида.
Фагоцитоз . Крупные молекулы органических веществ, например белков и полисахаридов, частицы пищи, бактерии поступают в клетку путем фагоцита (греч. "фагео" - пожирать). В фагоците непосредственное участие принимает плазматическая мембрана. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в "мембранной упаковке" погружается внутрь клетки. Образуется пищеварительная вакуоль и в ней перевариваются поступившие в клетку органические вещества.
Объясняет эта теория и различия метаболизма цитоплазмы и органоидов. Анаэроб - протоэукариот приобрел бактерий, которые уже стали аэробными (митохондрии); гетеротроф приобрел фототрофов (хлоропласты).
Теория симбиогенеза предсказывает гомологию (сходство) последовательностей ДНК органелл и бактерий. Это предсказание с появлением методов секвенирования блестяще подтвердилось. Например, по нуклеотидным последовательностям 16S-рибосомальной РНК хлоропласты наиболее близки к цианобактериям, а митохондрии - к пурпурным бактериям. И те, и другие рРНК резко отличаются от рРНК эукариотических рибосом цитоплазмы хозяев. Наконец, теория симбиогенеза предсказывает возможность множественного (неоднократного) приобретения симбионтов и вероятность нахождения нескольких разных свободноживущих бактерий, похожих на их предков. Видимо, эта возможность была реализована в случае хлоропластов.
Здесь надо отметить, что в большинстве книг по цитологии ограничиваются описанием хлоропластов зеленых растений: их-то мы себе и представляем, когда речь идет про эти органоиды. Сходное строение имеют хлоропласты зеленых водорослей. А вот у других водорослей они могут существенно отличаться как по строению мембранных частей, так и по набору пигментов.
У красных водорослей хлоропласты содержат хлорофилл, а и фикобилины - белковые пигменты, собранные в особые тельца - фикобилисомы; мембранные мешочки - ламеллы - расположены в них поодиночке. По этим признакам они наиболее (из всех хлоропластов) схожи с цианобактериями, прямыми потомками которых, видимо, и являются.
У зеленых водорослей и высших растений есть хлорофиллы a и b; нет фикобилинов; ламеллы собраны в стопки - граны. От типичных цианобактерий они по этим признакам отличаются. И вот в 70-е г.г. ХХ века был подробно описан замечательный прокариот-фотоавтотроф - Prochloron. Он был известен и ранее - это симбионт асцидий-дидемнид, у которых он живет не внутри клеток, а в клоакальной полости. (Дидемниды - мешковидные, прозрачные колониальные существа, обитающие на коралловых рифах. Из-за симбионтов имеют ярко-зеленую окраску. Могут медленно двигаться, выбирая освещенные участки. Симбионты получают от хозяина защиту и необходимые им ростовые вещества, а в ответ делятся с ним продуктами фотосинтеза и, видимо, аминокислотами - прохлорон способен к азотфиксации.) Оказалось, что прохлорон - скорее всего, цианобактерия (хотя некоторые ученые выделяют его в особый отдел Prochlorophyta вместе с позднее открытой свободноживущей нитчатой бактерией Prochlorothryx), но... у него нет фикобилинов; есть хлорофиллы a, b; есть стопки ламелл. Таким образом, это - "модель" предка хлоропластов высших растений!
По материалам Интернета.